Une nouvelle théorie pour l’évolution des chromosomes sexuels
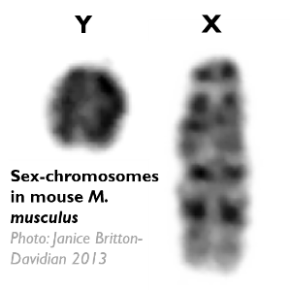
Chez de nombreuses espèces, le déterminisme du sexe est chromosomique. Par exemple chez les mammifères, les mâles sont XY et les femelles XX. Le sexe mâle est le sexe hétérogamétique (c’est-à-dire produisant des gamètes différents, portant soit un X soit un Y). Chez les oiseaux et les papillons, en revanche, c’est l’inverse, les femelles sont hétérogamétiques (ZW) et les mâles homogamétiques (ZZ). Comment se forment ces chromosomes sexuels et comment évoluent-ils ?
On observe que dans de nombreuses espèces, ces chromosomes cessent de recombiner entre eux. Ce phénomène d’arrêt de recombinaison survient souvent par « strates » successives que l’on peut documenter en observant différents paliers dans la divergence génétique entre ces chromosomes. On observe également souvent que le Y (ou le W) est dégénéré, c’est-à-dire qu’il accumule des mutations délétères au point de perdre la plupart des gènes dans sa partie non-recombinante. Finalement lorsque le Y (ou le W) est dégénéré, on observe fréquemment une compensation de dosage, c’est-à-dire que l’expression des gènes du X (ou du Z) est doublée dans le sexe hétérogamétique, ce qui compense cette perte de gènes.
Identifier les raisons évolutives menant à ces particularités des chromosomes sexuels est une préoccupation presque aussi vieille que leur découverte (en 1905, réalisée indépendamment par Stevens et Wilson). Au-delà de cette problématique, et des maladies génétiques associées au sexe comme l’hémophilie, l’étude des chromosomes sexuels a toujours eu une importance plus générale. Elle permet en effet d’étudier les causes et les conséquences de l’arrêt de recombinaison, et donc, en creux, les raisons qui pourraient favoriser le maintien de la reproduction sexuée (dont la recombinaison est un aspect central) dans la grande majorité des espèces.
La théorie actuelle « classique » expliquant la formation des chromosomes sexuels a été affinée pendant près de 50 ans. Elle comprend trois étapes qui s’enchaînent. La première étape est l’arrêt de recombinaison. L'idée prédominante pour expliquer celle-ci est qu'elle permettrait d’associer de manière permanente au Y des allèles favorables pour les mâles mais défavorables pour les femelles (le même argument inversé fonctionne pour le cas ZZ/ZW). Ces allèles à effet sexe-antagoniste sont ceux contribuant au dimorphisme sexuel. La deuxième étape, l’accumulation de mutations délétères sur le Y ou le W (la « dégénérescence ») survient car, en absence de recombinaison, la sélection est bien moins efficace à éliminer les mutations délétères des populations. Ce phénomène, appelé « interférence sélective », a été très étudié du point de vue théorique. La troisième étape, la compensation de dosage, correspond à l’évolution de régulateurs de l’expression des gènes pour rétablir un niveau d’expression adéquat des gènes qui ont dégénéré sur le Y ou le W (une surexpression de la copie du gène fonctionnelle qui reste sur le X ou le Z est favorisée car elle permet de rétablir un meilleur dosage des protéines dans la cellule). De nombreuses observations empiriques vont dans le sens de cette théorie classique: par exemple, il est clair que l’absence de recombinaison réduit l’efficacité de la sélection naturelle. Pourtant, l’ordre précis des processus et les relations exactes de cause à effet restent difficiles à prouver. C’est aussi l’absence d’alternative crédible qui a conforté cette théorie.
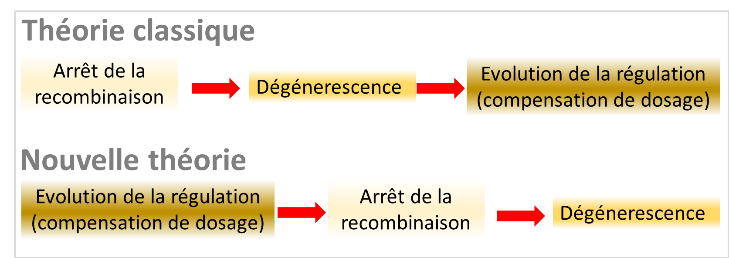
Dans un nouvel article publié dans Science, Lenormand & Roze proposent une théorie alternative, reposant sur un enchaînement des événements et une causalité différente. Au moyen de simulations informatiques, les auteurs montrent que cette nouvelle théorie explique mieux certaines caractéristiques de l’évolution des chromosomes sexuels que la théorie classique: en particulier, un arrêt progressif de la recombinaison le long du chromosome Y (ou W), et l'évolution rapide de la compensation de dosage. L’inversion de causalité repose sur l'idée que des arrêts de recombinaison fortuits sur le Y (ou le W) peuvent mener à l'émergence précoce d’un début de compensation de dosage, entraînant ensuite la dégénérescence. C’est ce début de compensation de dosage qui crée des effets sexe-antagonistes, et maintient l’arrêt de la recombinaison sur le long terme. Ici, il n’y a donc pas besoin de supposer l’existence préalable de gènes à effet sexe-antagonistes sur les chromosomes sexuels. Le dimorphisme sexuel n’aurait donc pas de lien causal direct avec l’évolution des chromosomes sexuels.
En renouvelant entièrement notre vision des scenarios possibles d’évolution des chromosomes sexuels, cette nouvelle théorie ouvre un vaste champ d’expérimentations et de tests empiriques. Elle montre également l'importance de prendre en compte les mécanismes de régulation de l’expression des gènes dans la théorie de l'évolution.
Laboratoires CNRS impliqués
- Centre d'Écologie Fonctionnelle et Évolutive CEFE (CNRS / EPHE / IRD / Université de Montpellier)
- Laboratoire Biologie Évolutive et Écologie des Algues EBEA (CNRS / Sorbonne Université / Station Biologique de Roscoff)
Référence
Lenormand T, Roze D. 2022. Y recombination arrest and degeneration in the absence of sexual dimorphism. Science.